Rosnatky - rod plný zvláštností
Nejznámější a nejhojnější z 11 druhů masožravých rostlin v české květeně je rosnatka okrouhlolistá (Drosera rotundifolia L.). Vyniká obrovským cirkumpolárním areálem a je nejběžnějším druhem rosnatky na severní polokouli. Sotva však může být pokládána za skutečně charakteristickou reprezentantku rodu, když většina z jeho asi 130 druhů roste na opačné polokouli a ve zcela jiných biomech. Jak různé jsou tyto biomy, tak se liší vzhled příslušných rosnatek.
Největší bohatství druhů i životních forem se nalézá v Austrálii. Četné tamější druhy a poddruhy jsou nově popsané, přičemž mají jen provizorní jména, nezveřejněná dosud platně podle závazných pravidel.

Rosnatka Burmannova (Drosera burmannii) mívá tak rychlý pohyb okrajových tentakul, že jej lze sledovat během 1 minuty. Tato atraktivní rosnatka je krátkověká a musí se proto stále obnovovat z výsevu. Je však samosprašná a semena spolehlivě klíčí.
Málokterý druh rosnatky má rozsáhlý areál, většinou jsou to endemity malých území. Jen několik druhů je dostatečně dostupným materiálem pro fyziologický nebo biochemický výzkum. V budoucnu proto není vyloučen větší význam ve farmacii. Možné je snad i biotechnologické využití, neboť rosnatky mají schopnost zvláště výborně vegetovat v definovaném prostředí laboratorních kultur "in vitro". Po biochemické stránce nejsou ovšem jednotnou skupinou. Mohou obsahovat jeden i více naftochinonů, z nichž nejčastější jsou 7-metyljuglon a plumbagin. Různé extrakty z rosnatek se používaly nebo ještě používají vně i vnitřně, a to nejen na kašel (D. filiformis, D. intermedia, D. ramentacea, D. rotundifolia aj.). Působí prý proti bradavicím (D. indica, D. rotundifolia), mohou být součástí infúzí proti arterioskleróze, v Indii se uplatňovaly proti syfilis (D. peltata), v Americe zase jako afrodisiakum (D. anglica, D. filiformis). Přehnané používání může vyvolávat záněty, což se ví o několika druzích včetně D. rotundifolia.

Africká Drosera collinsiae je patrně hybridního původu; dobře se rozsemeňuje a je vhodná pro větší rozšíření v kultuře.
Biochemickým otázkám byly věnovány i četné pokusy slavného C. Darwina s rosnatkou okrouhlolistou. Toho ovšem zajímaly hlavně enzymy a funkce trávicího aparátu. Lepkavé listy rosnatek totiž nejsou nijak jednoduchým zařízením.
Lapání a trávení kořisti
Rosnatky jsou hmyzožravé v pravém slova smyslu a svou kořist lákají pomocí krůpějí sekretu na koncích mnoha stopkatých žláz. Význam má snad i rudé barvivo droserin, obsažené často ve žlázách. Listy jsou potom třpytivé i barevné. Zdá se, že nějaký pach působí i na chemoreceptory hmyzu.
Potřísníme-li si výměškem prsty, což se nezřídka dokonale splní při přesazování robustních druhů rosnatek, přesvědčíme se o jeho slizké konzistenci a lepkavosti. Nakonec ještě zjistíme, že potřísněné ruce neočistíme pouhým proudem vody, mýdlem pak s obtížemi. Při drobném poranění se navíc uplatní přítomné enzymy a rána pálí. Podobným účinkům jsou jistě vystaveny citlivé tkáně kořisti, jíž vysoce smáčivý sekret zahlcuje dýchací štěrbiny na zadečku. Právě proto můžeme zprvu zaznamenat zmatek a neúčelné chování hmyzu po dosednutí na list rosnatky, což je důležité pro lovecký úspěch rostliny. Kořist postupně umdlévá, avšak zůstává dlouho živá, vystavena účinkům stále silněji vylučované trávicí tekutiny.
Síla hmyzu je všeobecně známá a mnohá kořist rosnatkám uniká. Naději na úspěch zvyšují rosnatky třemi způsoby. Jedním z nich je velmi protažený tvar čepele, tvořící dlouhou dráhu pro pohyb postupně slábnoucí kořisti. Druhým způsobem je růst v kobercích, vzniklých rozséváním semen z rozkývaného stvolu (D. aliciae aj.), tvorbou rostlinek z adventivních pupenů na starých listech (D. capillaris), anebo ze speciálních rozmnožovacích tělísek, odlamovaných a šířených deštěm (D. pulchella aj.). Kořist unikající z jedné rostliny se dostává vzápětí na další, přičemž se vysiluje. Třetím způsobem ke zlepšení loveckého úspěchu je vytváření spleti drobně olistěných prýtů, v níž může uvíznout i větší kořist, například motýl (D. stolonifera, D. macracantha a další.).
Lapací orgány rosnatek nejsou účinné na každý hmyz. Především velikost kořisti, její síla a snad i míra ochlupení a etologie jsou faktory určující, zda se příslušný živočich může, nebo nemůže stát obětí určitého druhu rosnatky. Nedivme se proto, že rosnatka neuchvátí ani svou obvyklou přirozenou kořist, narušíme-li hmyzí chování při umělém krmení. Rosnatky také mohou být napadeny mšicemi, jež vyhledávají nežláznaté části a nikdy nejsou přilákány žlázami. Housenka pernatušky Trichoptilus paludum, žijící i v Čechách, je odolná vůči sekretu rosnatky okrouhlolisté. Okusuje listy, maskována pomocí mimikry. Z jihozápadní Austrálie jsou známy bezkřídlé ploštice rodu Cyrtopeltis, pohybující se po listech rosnatek, přiživujíce se na přilepené kořisti.
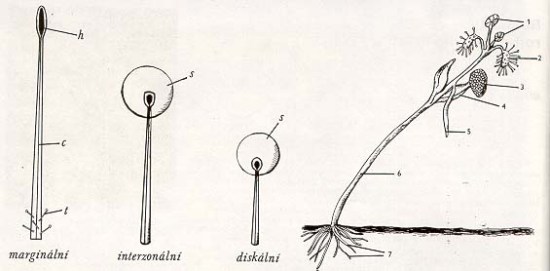
Vlevo tři typy tentakul rosnatky Burmannovy (stálá kapka sekretu u diskálních a interzonálních tentakul vždy chybí u marginálních, jejichž hlavičky jsou i bez sekretu silně přilnavé: h - hlavička c - céva ve stopce žlázy, t - trichomy (chlupy), s - sekret. Vpravo: zvláštní způsob klíčení rosnatek (D. cistiflora); celková velikost 5 mm, 1 - vyvíjející se listy, 2 - první lapací listy, 3 - osemení, 4 - děložní lístky, 5 - první adventivní kořen spouštěný do půdy, 6 - článek podděložní, 7 - kořenové vlášení na redukovaném a později potlačeném primárním kořeni.
Žlázy rosnatek se často označují termínem "tentakula" (jedn. č. tentakulum tj. chapadélko). Jsou to výčnělky (nikoli tedy chlupy), napojené na cévní systém rostliny, zakončené hlavičkou vylučující sekret. V něm byly zjištěny mukopolysacharidy způsobující slizkost, organické kyseliny a enzymy, schopné rozkládat svalovinu, tuky i některé další látky, ne však chitinové obrnění hmyzu. Vlivem přítomnosti kořisti na listu se zvyšuje sekrece. Dráždivě přitom působí jednak pohyb hmyzu, jednak prvé produkty jeho natrávení zaživa. Mechanický i chemický vzruch způsobuje změnu původního, klidového elektrochemického potenciálu v listu. Takový signál se potom šíří pletivy rychlostí asi 5 mm za sekundu z místa podráždění do okolí. Je dobře měřitelný, podobně jako u živočišných nervů, neboť má dosti značnou velikost, kolem 60 mV.
Skutečně nápadnou reakcí je pomalé ohýbání stopek žláz. U řady druhů rosnatek (avšak ne u všech) následuje i ohýbání listu tak, aby se zdokonalil kontakt s kořistí. Pohyb listu je růstový, působený rozdílnou rychlostí růstu svrchní a spodní plochy čepele. Tento děj je řízen vlivem auxinu, fytohormonu, jenž je produkován mladým pletivem na konci listu a díky elektrické polaritě jeho molekul je usměrňován do oblasti dráždění.
Zajímavější je ovšem činnost samotných žláz. Jejich pohyby bývají u většiny druhů zjevné asi po 20 minutách od uvíznutí kořisti na lepkavém listu. Obzvláště brzkými a rychlými pohyby je známa tropická rosnatka Drosera burmannii. Při optimální teplotě kolem 30 °C (nízká teplota reakce tlumí) lze ohýbání žláz jasně vidět již během jediné minuty. Dosud se asi neví, že ještě rychlejší může být zřídka pěstovaná jihoafrická D. pauciflora.
Žlázy na ploše listů rosnatek bývají kratší než na obvodu. Lišit se mohou i tvarem hlaviček. Nejdůležitější rozdíl však spočívá ve funkcích. Obvodová, čili marginální tentakula slouží k posunutí ulpělé kořisti do ideální polohy. Reagují nejsilněji na mechanické dráždění a ohýbají se vždy stejně, jen směrem ke středu či ose listu. Je to pohyb, který je přítomností kořisti sice vyprovokován, ale nikoli usměrňován (případ tzv. nastie). Tentakula na ploše listu, označovaná též "diskální", reagují spíše na chemické dráždění. Ohýbají se kterýmkoli směrem, orientovaně ke kořisti (případ tzv. tropismu).
U některých rosnatek nejsou rozdíly mezi marginálními a diskálními žlázami příliš vyhraněné. Nejčastěji jsou však velmi zjevné. Někdy se morfologicky odlišují ještě tzv. tentakula interzonální, jež se pozicí i funkcí nacházejí na přechodu mezi předešlými. U atraktivní australské rosnatky D. binata jsou nápadná svou délkou, takže přesahují i tentakula marginální.
Trávení kořisti trvá několik dní a tato doba závisí na kondici rostliny, druhu kořisti a na teplotě. Nezbytné jsou přitom symbiotické bakterie a houby, což je běžné u masožravých rostlin, ale i u živočichů. Po vstřebání živin se list vyrovnává do původního tvaru a z kořisti zbývá prázdný chitinový skelet. Lapací schopnost listu klesá vlivem rychlého stárnutí, v důsledku opotřebení neustále vyměšujících žláz. Nejstarší listy, neschopné již lapání, slouží ještě nějaký čas jako asimilační fotosyntetické orgány.
Právě brzké opotřebovávání lapacích listů souvisí s charakteristickou vlastností rostlin s adhezními pastmi, kterou je rychlá obměna listů. Rapidní růst ovšem nemůže být plně hrazen z kořisti, neboť ta je především zdrojem nedostatkových biogenních prvků. Neméně důležitý je i patřičný příkon energie, a proto jsou rosnatky velmi závislé na vydatném oslunění. I druhy s minimálními požadavky (kuriózní a nepočetné případy adaptované na růst v tropickém deštném lese), prosperují nejlépe při vydatném, byť rozptýleném osvětlení. V přírodě je proto i pro ně světlo limitujícím ekologickým faktorem.
Permanentně rostoucí druhy
Mnohotvárný a ekologicky rozrůzněný rod Drosera je třeba pro toto pojednání rozčlenit, i když to je možné jen nedokonalým způsobem. Jednu výraznou skupinu tvoří druhy z celoročně natolik teplých i vlhkých stanovišť, že rostliny netvoří klidová neboli dormantní stadia. Právě takové jsou i nejčastěji pěstované rosnatky D. capensis, D. capillaris a mnoho dalších.

Rosnatka kopisťovitá (D. spathulata) je velmi variabilní druh pocházející z východní Asie, Austrálie a Nového Zélandu. Snímky M. Studničky.
V popisované skupině jsou druhy někdy značně podobné, a to i navzdory výskytu na různých kontinentech. Chyby v pojmenování jsou proto časté a semena mylně určených druhů šíří i některé botanické zahrady. Druh D. aliciae se například zejména dříve označoval jako D. spathulata. Skutečná D. spathulata je sama o sobě dosti variabilní, a tak se pěstuje v různých kultivarech se vžitými, avšak platně nepublikovanými jmény. Pod falešným označením dnes ovšem nejčastěji obdržíte americkou D. capillaris, odolnou a samovolně se rozsemeňující rosnatku. Aby bylo možné odstranit alespoň problémy jmen nejvíce frekventovaných v inventářích sbírek, sestavil jsem klíč k určování vybraných druhů. V něm je zcela vynechán běžný, avšak příliš nepřesný termín "kopisťovité listy" a je nahrazen výrazy praktičtějšími (viz obr.).
Určovací klíč
A - listy lžicovitého tvaru (typu D. capillaris)
1a) Řapík klínovitý, uprostřed alespoň 2 mm široký: 2
1b) Řapík čárkovitý, jen asi 1 mm široký: 3
2a) Růžice ploché, přiléhající k půdě: D. dielsiana Exc. et Laund.
2b) Růžice chocholovité: D. venusta Debbert
3a) Řapíky krátké, většinou do 10 mm, kališní lístky ostře špičaté: D. communis St. Hil.
3b) Řapíky dlouhé, zpravidla více než 10 mm, kališní lístky tupé: 4
4a) Květenství jen 4-6 květá, stopečky květů a kalichy žláznaté, dřípené palisty jen 2-3 mm dlouhé: D. burkeana Planch.
4b) Květenství mnohokvětá (více než 6), stopečky květů a kalichy lysé, dřípené palisty 3-5 mm dlouhé: D. capillaris Poir.
B - listy klínovité (typu D. natalensis)
1a) Průměr růžice pod 40 mm: 5
1b) Průměr růžice nad 40 mm: 2
2a) Listy v polovině zaškrcené (houslovitý tvar), přičemž dolní část je zcela beze žláz: D. slackii Cheek
2b) Listy nezaškrcené, žlázy plynule řídnou směrem k bázi: 3
3a) Čnělky dole srostlé ve sloupek: D. hamiltonii C. R. P. Andr.
3b) Čnělky volné: 4
4a) Čnělky jedenkrát, a to dole vidličnatě rozvětvené, semena oválná: D. cuneifolia L. F.
4b) Čnělky větvené alespoň dvakrát, a to jednou dole, podruhé výše, semena úzce vřetenovitá: D. natalensis agg.*)
5a) Květenství vysoká do 8 cm, palisty redukované ve 2 chlupovité výčnělky (fimbrie), anebo zcela zaniklé: D. brevifolia Pursh
5b) Květenství vyšší než 8 cm, palisty řádně vyvinuté a dřípené v několik cípů: 6
6a) Znělky nejširší na bázi, semena oválná: D. spathulata Labill.
6b) Čnělky nejširší na konci, semena vřetenovitá: D. montana St. Hil.
*) D. natalensis agg. značí agregát v kultuře sotva rozlišitelných a určitě blízce příbuzných mikrospecií. Kromě D. natalensis Diels jsou to D. aliciae Hamet a několik dalších nověji popsaných [Debbert, Mitt. Bot. Staatsamt. München 23 (1987): 431-436 a 30 (1991): 373-380].
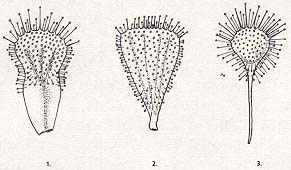
Různé tvary listů rosnatek: 1 - houslovitý (D. slackii), 2 - klínovitý (D. cuneifolia), 3-lžicovitý (D. capillaris). Kreslila R. Novotná.
Ekologie
Skupina zahrnuje rostliny otevřených formací, například rašelinných mokřadů a horských pramenišť v tropech, různých podmáčených savan a podobně. Jsou to rostliny světlomilné. Ve skleníkových podmínkách proto zpravidla strádají přes zimu, kdy je málo slunce a dny jsou mnohem kratší než v tropech a subtropech. Tehdy dochází k útlumu růstu, snížení sekrece a též poklesu odolnosti vůči patogenním plísním. Některé druhy nelze vůbec pěstovat bez preventivních postřiků fungicidy, jež naštěstí rosnatky dobře snášejí (D. cuneifolia).
Půdy osídlované rosnatkami bývají rašelinné, velmi chudé minerálními živinami, často silně písčité, nevápnité a silně kyselé (pH 3,5 až 4,5). Někdy rostliny koření i v kobercích mechů a rašeliníků. V kultuře se používá rašelina smíšená s hrubým bílým křemenným pískem, anebo rašelina bez příměsí. Poměr písku a rašeliny má vliv na fyzikální vlastnosti směsi, ale ty také ovlivňuje kvalita samotné rašeliny. Světleji hnědá, vláknitá nebo vrstevnatá bývá pro rosnatky příznivější než kávové hnědá, s blátivým charakterem. Půda má být prostupná pro kořeny, vodu i plyny.
Osázené nádoby mají stát celoročně nejméně do 2/3 výšky ve vodě. Čas od času je správné vodu vypustit a za den či dva opět napustit. Tím docílený vertikální pohyb vody v půdě vypuzuje plyny vznikající při rozkladu humusu. Mezi nimi jsou i nenasycené uhlovodíky, účinkující jako inhibitory růstu. Jsou obvykle příčinou nápadného zhoršování kondice rosnatek v kultuře. Nebývá to tedy často podezřívaný vápník ze zálivky, neboť ten je fytotoxický až při posunu půdní kyselosti blízko k neutralitě. Při vysoké pufrační kapacitě rašeliny k tomu v praxi nedochází.

Drosera admirabilis
Někteří pěstitelé chválí výsledky mírného hnojení rosnatek postřikem nebo zálivkou velmi zředěnými roztoky minerálních hnojiv. To souhlasí s terénními poznatky, že například D. rotundifolia na mírně úživných slatinách mívá neobyčejně mohutný vzrůst. Trofické optimum rosnatek je však přitom velmi nízké a úzké, takže zprvu mnohoslibné experimenty s přihnojováním jsou provázeny rizikem zničení rostlin.
Důležité jsou i teploty, zejména v zimě. Snížení na 12 - 16°C, pro řadu subtropických druhů zcela přirozené, napomáhá určitému vegetačnímu útlumu. Díky tomu rosnatky netvoří v období nedostatku světla "nevyzrálá" pletiva, náchylná k houbovým infekcím. Mykózám je třeba čelit také dobrým větráním. Rosnatky z popisované skupiny se proto, až na nepočetné výjimky, nehodí pro uzavřené pokojové vitríny.
Generativní a vegetativní rozmnožování
Semena rosnatek klíčí vždy na povrchu substrátu, vystavena světlu. Bývají velmi malá, až práškovitá. Jejich osemení je buď síťovitě ztenčené a kolem semene volné (anemochorní druhy), anebo díky silnému zvrásnění dvůrkaté (hydrochorní druhy). Nepatrné rozměry semen umožňují vznik jen miniaturních, anatomicky velmi zjednodušených semenáčků. Ač mají 2 dělohy, článek podděložní (hypokotyl) a kořenové vlášení jako pozůstatek po redukovaném primárním kořenu, v botanické literatuře se označují pro tento případ nesprávným termínem "prokaulomy" . Svádí k tomu způsob dalšího vývinu, kdy z vrcholu semenáčku v době tvorby prvních lapacích listů raší i adventivní kořen. Dolní část rostlinky potom zaniká a tento kořen ukotví mladou rosnatku v substrátu. Semenáček ve stadiu před tvorbou lapacích listů je tedy jen provizorním zařízením.

Drosera slackii
Pro druhy osídlující jen dočasně příznivé biotopy jsou semena důležitou "pojistkou" proti vymření v důsledku vytlačování konkurujícími druhy. Semena slouží k úniku na větší vzdálenosti a jejich velký počet poskytuje naději na dosažení nového příležitostného stanoviště. Rosnatky s touto životní strategií bývají přímo samosprašné, a tudíž nezávislé na opylovačích a přízni počasí v době květu. Příkladů je mnoho (D. aliciae, D. brevifolia, D. capillaris aj.).
Nepohlavní rozmnožování u popisné skupiny není běžné, i když některé druhy tvoří dceřiné rostliny z adventivních pupenů na starých listech, doléhajících na půdu (D. capillaris, D. roraimensis aj.). Rosnatky jsou však příznačné pro různě rozrušované (disturbované) biotopy, a mají proto dobrou regenerační schopnost při poškození. Toho lze využívat při umělém vegetativním množení.
U druhů se silnými kořeny lze získat mladé rostliny z asi 2 cm dlouhých segmentů, zatlačených vodorovně do povrchu mokré rašeliny. Také odtržené listy, přitlačené na povrch rašeliny a řídce posypané rozstříhaným živým rašeliníkem pro snížení výparu, mohou vyrašit v mladé rostlinky.
Copyright © Miloslav Studnička, 1993